











Smo na morju v bližini Pirana, na vzorčevalni postaji z oznako 000F, ki leži nedaleč od oceanografske boje Vida. Piše se trinajsti december, ura pa je deset zjutraj. Dežuje. Milijan, tehnik z Morske biološke postaje Piran (NIB-MBP), v morje spušča Niskinove vzorčevalnike, pričvrščene na vzorčevalno rozeto. Njegova naloga je zbrati vzorce morske vode z različnih globin in s sondo izmeriti pomembne parametre, kot so slanost, temperatura in koncentracija raztopljenega kisika. To počne že dobrih trideset let, enkrat mesečno na istem mestu. Nekaj litrov vode nekaj kilometrov od obale. Kaj išče? Zakaj si na Morski biološki postaji Piran vsa ta leta prizadevajo, da ostaja vzorčenje prav na tej postaji aktivno? In kaj je Niskinov vzorčevalnik?
Ko je morje ustekleničeno in pripeljano v laboratorij, je na pogled vedno enaka prozorna tekočina, a videz vara. Voda je namreč glede na fizikalne, kemične in biološke lastnosti lahko zelo različna. Morje pred Piranom je (kot v celotnem Tržaškem zalivu) namreč zelo dinamično. Tu se voda, ki iz Levantinskega morja na najbolj vzhodnem delu Sredozemlja pripotuje ob vzhodni jadranski obali in se na svoji poti že dodobra spremeni, obogati s številnimi pritoki vodotokov. Ker je morje plitvo, je dovzetno za močne vplive zunanjih dejavnikov, kot so sonce, veter in drugi vremenski pojavi, ti pa neenakomerno vplivajo na različne sloje morske vode. Na tem mestu imamo že prvi odgovor: Niskinov vzorčevalnik je pripomoček, ki omogoča vzorčenje vode na točno določeni globini, ne da bi se vzorec med dvigom pomešal z drugo vodo.
Vzorci, ki jih vsak mesec pripeljejo v laboratorij, v sebi skrivajo tudi bogato biotsko pestrost. V vsakem litru vode živi izjemno veliko organizmov, ki so mikroskopskih velikosti in zato prostemu očesu praktično nevidni. V dolgoletnem nizu vzorčenj nas med drugim zanima skupina sicer zelo raznolikih organizmov, ki jim pravimo fitoplankton. To so planktonski organizmi, ki niso aktivni plavalci in so večinoma avtotrofni, torej sposobni fotosinteze. K fitoplanktonu prištevamo še številne druge organizme z zelo raznolikimi metabolizmi, poleg avtotrofov tudi miksotrofe in celo sorodne heterotrofne organizme. Njihova ekosistemska vloga v morjih in oceanih je podobna vlogi rastlin na travnikih in v gozdovih, saj predstavljajo enega od stebrov morske prehranjevalne mreže in so nepogrešljiv člen biogeokemičnega kroženja.
Za fitoplankton je značilna velika raznolikost oblik in tudi velikosti, saj lahko celice merijo od okoli 2 µm do več kot 200 µm, vendar prepoznavanje vrst pod mikroskopom kljub temu ni enostavno. Nekatere značilnosti pod svetlobnim mikroskopom niso vidne, poleg tega so si nekatere vrste morfološko tudi zelo podobne. Zato potrebuje strokovnjak za fitoplankton dolgoletne izkušnje in nenehno učenje, kar zahteva veliko časa in energije. Če bi torej želeli prepoznavati prostorsko variabilnost fitoplanktona v morju, ki je lahko glede na raznolike značilnosti vodnih mas precejšnja že na manjšem območju, kot je denimo Tržaški zaliv, bi potrebovali kopico strokovnjakov, ki bi lahko pregledali veliko število vzorcev. Ta vidik smo večinoma prepustili satelitom, ki spremljajo barvo oceanov in nam podajajo informacijo o biomasi, v zadnjem času pa tudi o sestavi fitoplanktona v površinskem sloju morij in oceanov. Ker pa satelitski podatki še zdaleč niso tako natančni kot rezultati analize v laboratoriju, smo v zadnjih letih svoje raziskovanje usmerili v prepoznavanje časovnih vzorcev fitoplanktona, tudi v povezavi s podnebnimi spremembami in drugimi vplivi človeških dejavnosti. Tu torej tiči odgovor na še eno vprašanje iz uvoda: cilj večdesetletnega vzorčenja morske vode na postaji 000F je identificirati sestavo fitoplanktonske združbe, razložiti njene ekološke značilnosti in ugotoviti, kako se spreminja v času.
Preden se lotimo opisa raziskave (njene rezultate smo nedavno objavili v znanstveni reviji Science of the Total Environment), je nujno kratko teoretično pojasnilo. Že pionirji ekološke vede so razmišljali o lastnostih in omejitvah ekološke teorije in zakonitosti. Eden od konceptov, ki je prav pri fitoplanktonu naletel na protislovna spoznanja, je koncept ekološke niše. Ekološka niša zajema prostor, ki ga zaseda neka vrsta, vlogo te vrste v njem ter njegove biotske in abiotske lastnosti (npr. temperaturo, slanost, prisotnost plenilcev). Iz tega sledi, da je mogoče okoljske značilnosti uporabiti za napovedovanje prisotnosti nekega organizma. Poleg tega naj bi določen prostor zaradi načela kompetitivne izključitve lahko zasedalo le omejeno število vrst, vendar pa to pri fitoplanktonu ne drži, zato se je uveljavil izraz planktonski paradoks.
Teoretično je namreč ekoloških niš, primernih za fitoplankton, v morskem okolju malo, poleg tega so tudi kratkožive. A v praksi je sobivajočih vrst kljub temu izjemno veliko. V preteklosti je bilo ponujenih veliko razlag za to paradoksalno bogastvo vrst, pred kratkim pa je teoretični razvoj prinesel nov preobrat. Ekologi so, izhajajoč iz nekaterih starih dognanj, oblikovali dve med seboj povezani teoriji: nevtralno teorijo (angl. neutral theory) in teorijo grudičastega sobivanja (angl. lumpy coexistence theory). Nevtralna teorija zavrača koncept ekološke niše in do neke mere tudi koncept populacije organizmov, saj obravnava vsak organizem kot samostojno bitje, čigar možnosti za preživetje so individualne in plod naključja. Čeprav je ta teorija zelo poenostavljena, se je v nekaterih primerih izkazala za dobro utemeljeno, njen uspeh pa je nato preko povezovanja s konceptom ekološke niše pripeljal do teorije grudičastega sobivanja. Ta pravi, da je uspeh neke vrste znotraj ekološke niše odvisen od niza naključnih usod posameznih osebkov. Številne raziskave ekologije fitoplanktona so neodvisno od opisanega teoretičnega razvoja privedle do preseganja koncepta ekološke niše za posamezno vrsto. Vse pogosteje se preučuje fitoplanktonske združbe (angl. assemblages), torej skupnosti različnih vrst v določenem vodnem telesu v določenem času. Pri naših raziskavah smo izkoristili ta teoretični napredek in tudi dobljeni rezultati so v skladu s pričakovanji opisanih teorij.
V tridesetih letih spremljanja fitoplanktona na postaji 000F smo opazili več kot 150 vrst, ob tem pa tudi precejšnje število organizmov, ki nam jim s svetlobnim mikroskopom ni uspelo določiti vrste in jih uvrščamo v višje taksonomske skupine. Prvi korak v vrsti analiz teh podatkov je bil osredotočen na iskanje ponavljajočega se vzorca v vrstni sestavi. Na podlagi številnosti in soprisotnosti v mesečnih vzorcih morske vode smo fitoplanktonske vrste, ki spadajo v različne taksonomske skupine, s pomočjo hierarhičnega razvrščanja in nekaterih drugih metod združili v skupine – združbe. Za vsako od tako identificiranih združb smo lahko opisali sezonsko in medletno dinamiko, saj so se med seboj močno razlikovale po trajanju in cikličnosti obstoja: nekatere so bile bolj kratkožive in so se v dolgoletni seriji pojavile le nekajkrat (ali celo samo enkrat), spet druge pa so pokazale svojo ciklično naravo in smo jih lahko okarakterizirali kot sezonske. Za vsako od fitoplanktonskih združb smo s posebnim indeksom identificirali značilne vrste. Na podlagi predhodnega poznavanja ekoloških preferenc teh vrst smo lahko oblikovali hipoteze o odnosu med združbami in okoljskimi dejavniki, ki vplivajo na njihovo pojavljanje.
Najlažje je bilo seveda poiskati preprosta linearna razmerja med pojavljanjem in številčnostjo določene vrste ali združbe ter posameznimi okoljskimi dejavniki, denimo temperaturo morja, slanostjo ali razslojenostjo vodnega stolpca. Vendar je to hkrati tudi pretirano poenostavljanje realnosti. Najpogosteje so namreč ta razmerja nelinearna in večfaktorska. Poglejmo na primer razslojenost vodnega stolpca, ki smo ga ovrednotili z gradientom gostote morske vode od površine do dna, in učinek te razslojenosti na točno določeno skupino organizmov, kokolitoforide (skupina bičkastih alg, ki imajo celice prekrite s kalcitnimi ploščicami različnih oblik). Njun odnos je pri nizkih vrednostih najprej negativen (kokolitoforidi z naraščajočo razslojenostjo upadajo), z naraščanjem gradienta gostote postane pozitiven (kokolitoforidi z naraščajočo razslojenostjo naraščajo) nato pa, ko gradient preseže določen prag, spet postane negativen (Slika 1). Pri razlagi tega primera je treba upoštevati tudi vzroke za razslojevanje vodnega stolpca, npr. daljše obdobje brez močnejšega vetra ali prisotnost površinskega sloja osladkane vode.
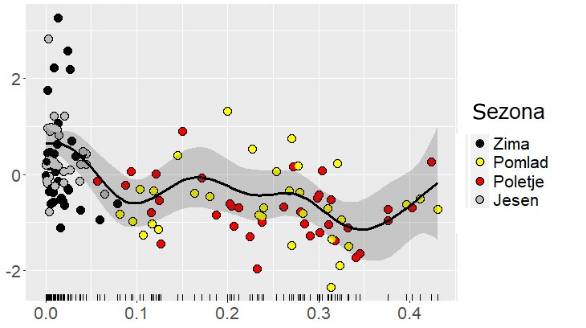
Slika 1: Posplošeni aditivni model (angl. generalized additive model – GAM) abundance fitoplanktonske skupine kokolitoforidov z gradientom razslojenosti vodnega stolpca kot kovariato
Zaradi nelinearnosti in prepletenosti različnih dejavnikov je vzpostavitev modela, ki bi dobro opisal odnos med okoljskimi dejavniki in fitoplanktonom, zelo zapletena, še bolj pa jo otežuje spreminjanje v času. Zato je zelo pomembno razumevanje dinamike okoljskih dejavnikov. Če to ponovno ponazorimo s primerom razmer v vodnem stolpcu: dlje časa trajajoči močnejši vetrovi, denimo burja, bodo spodbudili rast in razvoj tistih vrst, ki so dobro prilagojene na razmere v premešanem vodnem stolpcu (potrebujejo npr. visoke koncentracije hranilnih snovi in/ali turbulentno okolje, oboje pa je posledica mešanja vodnih slojev zaradi delovanja vetrov), posledično pa bodo te vrste prisotne tudi v mesecih kasnejšega zatišja in bodo lahko ob ugodnih razmerah spet prevladale. Poleg ciklično ponavljajočih se dogodkov so za meteorološke pojave značilne tudi epizode bolj ali manj ekstremnih razmer neperiodične narave, ki ravno tako vplivajo na fitoplanktonske združbe. Ugotovili smo, da je povprečna življenjska doba fitoplanktonskih združb (torej koliko časa neka združba vztraja, preden jo zamenja druga z drugačno sestavo) v Tržaškem zalivu 2–4 mesece, kar je tudi eden od značilnih tipov periodičnosti fitoplanktona v obalnih morjih in estuarijih (t. i. kratka periodičnost).
Zgornje ugotovitve in predpostavke so nas pri modeliranju odnosa med fitoplanktonskimi združbami in okoljskimi parametri vodile do uporabe nevronskih mrež. Nevronske mreže, ki so osnova za delovanje strojnega učenja, omogočajo napovedovanje nekega izida na podlagi množice podatkov. V našem primeru nas je zanimalo, kdaj in za kako dolgo se bo na nekem območju pojavila določena fitoplanktonska združba, če poznamo bolj ali manj kompleksen niz okoljskih parametrov, ki prav tako vplivajo drug na drugega. Poleg tega smo ta isti niz okoljskih podatkov ločili na ciklično in neciklično komponento ter analizirali, kako ti komponenti delujeta na modele nevronskih mrež. V raziskavi smo uporabili podatke o vetrovih (smeri in moči), padavinah, rečnih pritokih, slanosti ter temperaturi zraka in morja. Ker meteorološki in oceanografski pojavi praviloma delujejo na mezoskali (torej na območju z velikostjo 1–500 km), smo dobljene modele nevronskih mrež preizkusili tudi na podatkih iz Beneškega zaliva (s pomočjo soavtorjev z beneškega Inštituta za morske znanosti – ISMAR), ki je od naše postaje oddaljen približno 100 km. Ugotovili smo, da je s pomočjo okoljskih parametrov mogoče napovedati pojavljanje fitoplanktonskih združb v severnem Jadranskem morju in da periodični pojavi pomembneje prispevajo k uspešnosti modelov od neperiodičnih (primer jesensko-zimske združbe na Sliki 2).
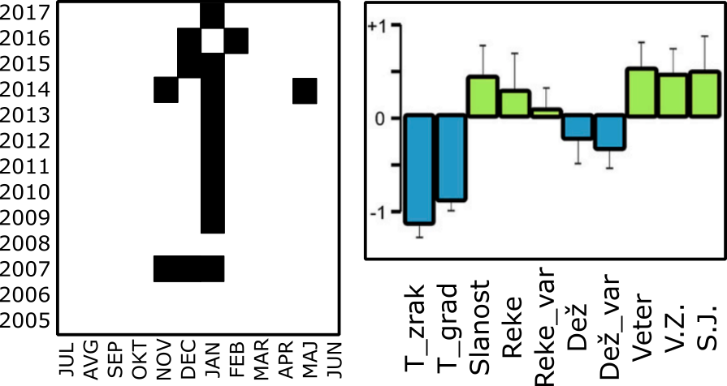
Slika 2: Primer časovne porazdelitve ene izmed fitoplanktonskih združb, za katero so bili značilni kokolitoforidi (levo), in okoljske značilnosti v času pojavljanja te združbe – pretežno jeseni in pozimi (desno). Okoljske značilnosti: temperatura zraka (T_zrak), temperaturni gradient med površino in dnom (T_grad), slanost morja (Slanost), pretok reke Soče (Reke), variabilnost pretoka reke Soče (Reke_var), padavine (Dež), variabilnost padavin (Dež_var), jakost vetra (Veter), prevladujoča smer vetra (V. Z., S. J.)
V jesenskem in zimskem obdobju je bila sukcesija fitoplanktonskih združb na dveh koncih severnega Jadranskega morja sinhronizirana, torej sta bili obe območji raziskav v tem oziru povezani, medtem ko so spomladi in poleti lokalno različne razmere botrovale obstoju različnih združb v obeh zalivih. Na obeh območjih je bilo mogoče najti fitoplanktonske združbe s podobnimi vrstami pod podobnimi okoljskimi pogoji, ne pa nujno ob istem času. Na primer: združba, za katero so značilne majhne diatomeje (tudi kremenaste alge, ekološko zelo pomembna skupina fitoplanktona s kremenasto celično steno) iz rodu Cyclotella (Slika 3), se razvije v pogojih toplega, razslojenega vodnega stolpca. Taki pogoji so bili v Tržaškem zalivu prepoznani pretežno spomladi, v Beneškem zalivu pa nekoliko kasneje poleti. Združbe, v katerih prevladujejo velike diatomeje in se v Tržaškem zalivu razvijejo ob močnejših jesenskih padavinah, so imele v Beneškem zalivu ravno tako drugačno dinamiko. Nasprotno so se v značilnih zimskih razmerah v obeh zalivih razvile združbe s tipičnimi zimskimi vrstami, ki pa so bile krajevno različne: majhna kozmopolitska vrsta kokolitoforidov Emiliania huxleyi (naslovna fotografija) v Tržaškem zalivu in diatomeje iz rodu Skeletonema v Beneškem zalivu. Vidimo torej zelo kompleksno sliko fitoplanktona v danem času in na danih (čeprav bližnjih) območjih: fitoplankton, ki ga raznašajo tokovi, se ob ustreznih pogojih hitro razmnožuje. Ker nenehno spreminja položaj, se tudi pogoji lahko hitro spremenijo, trenutno prisotnim vrstam pa lahko novi pogoji ustrezajo ali pa ne, kar botruje izbiri drugih vrst. Kljub temu pa cikličnost okoljskih dejavnikov zagotavlja do določene mere predvidljiv okvir, ki omogoča sukcesijo fitoplanktonskih združb.

Slika 3: Vrsta diatomej iz rodu Cyclotella, ki sestavlja pomladno ali poletno fitoplanktonsko združbo (foto: Petra Slavinec, posneto z elektronskim vrstičnim mikroskopom, pridobljenim v okviru projekta RI-SI-LifeWatch, MIZŠ in ESRR)
Izsledki raziskave predstavljajo pomemben korak v smeri razumevanja kompleksne ekologije fitoplanktona v obalnih območjih, pri tem pa so se časovno določene skupine sobivajočih vrst – združbe – izkazale kot dobro analitično orodje. Nadaljnjih raziskovalnih vprašanj seveda ne zmanjka, saj sestava fitoplanktonskih združb pomembno vpliva na druge procese v morju, denimo na razmere in spremembe v morskem prehranjevalnem spletu. Ker pa so vsi ti procesi odvisni tudi od podnebnih dejavnikov, v bližnji prihodnosti pričakujemo velike spremembe, ki se do neke mere kažejo že sedaj. Zaradi vse pogostejših ekstremnih dogodkov, kot so suše in poplave, se periodičnost nekaterih procesov v fitoplanktonu izgublja, kar vpliva na dogajanje v prehranjevalnem spletu in na produktivnost ekosistema. Da bomo spremembe lažje predvideli, tudi z nadaljnjo uporabo modeliranja, je zelo pomembna kontinuiteta vzorčenja fitoplanktona – vsak mesec nekaj litrov morske vode na isti vzorčevalni postaji.




