





- Deli
-

-

-

- Prenesi PDF
-

- Natisni
-

Predvajaj avdio zapis članka

Tokrat se pod kratico NLP ne skrivajo neznani leteči predmeti, temveč nekaj povsem drugega – Nep1 podobni proteini (angl. Nep1-Like Proteins). Poimenovanje Nep1 izhaja iz opažanja ob odkritju prvega proteina iz družine NLP, da ta ob stiku z rastlino spodbudi sproščanje rastlinskega hormona etilena in povzroči nekrozo rastlinskega tkiva (angl. Necrosis and Ethylene-inducing Peptide 1) (Bailey 1995). NLP‑je v naravi med svojimi napadi sproščajo številni rastlinski patogeni. Čeprav imajo vsi NLP-ji podobno zgradbo, nekateri izmed njih delujejo kot toksini, drugi pa ne – v prispevku se bomo osredotočili na prve, za katere je znano, da po »pristanku« na rastlinski celični membrani tvorijo majhne luknjice oziroma pore. Skozi pore lahko prehajajo ioni in majhne molekule, kar patogenu pomaga pri okužbi rastline. Zaradi sposobnosti tvorbe por NLP‑je uvrščamo med t. i. porotvorne proteine, pri čemer so še posebno zanimivi zato, ker predstavljajo edine znane porotvorne proteine, ki napadajo rastline. Poleg tega je njihov mehanizem tvorbe por povsem unikaten in še vedno poln ugank. Vemo, da so njihova tarča posebni lipidi, in sicer glikozil inozitol fosforil ceramidi oziroma lipidi GIPC (angl. Glycosyl Inositol Phosphoryl Ceramides). Ne vemo pa, zakaj nekatere rastline na NLP-je niso občutljive, čeprav vsebujejo lipide GIPC.
NLP-ji in zakaj jih preiskujemo
Kot smo že omenili, NLP-ji patogenom pomagajo pri napadu na rastlino, zaradi česar jih uvrščamo med efektorske proteine. Najdemo jih pri številnih med seboj nesorodnih rastlinskih patogenih iz vrst glivam podobnih organizmov oomicet, bakterij in gliv, ki so odgovorni za bolezni agronomsko pomembnih rastlin, kot so krompir, paradižnik, soja, vinska trta, kakavovec in tobak (Seidl in Van den Ackerveken 2019). Še danes povzročajo velike izgube pridelkov. NLP-ji so sodelovali tudi v zgodovinsko odmevni veliki lakoti, ki je prizadela Irsko med letoma 1845 in 1849, in sicer zaradi pojava krompirjeve plesni, katere povzročitelj je oomiceta Phytophthora infestans (Yoshida idr. 2013). Ta namreč pri napadu na rastlino uporablja tudi NLP-je (Haas idr. 2009).
NLP-ji so za raziskovanje zanimivi iz več razlogov, med drugim zato, ker so zaradi pojavnosti pri velikem številu patogenov potencialna tarča za razvoj (bio)pesticidov širokega spektra. Kot omenjeno, so edini znani porotvorni proteini, ki napadajo rastline, njihov mehanizem naluknjanja membrane pa je povsem drugačen od poznanega (Pirc idr. 2023). Drugi do sedaj znani porotvorni proteini namreč ob stiku z membrano spremenijo svojo obliko oziroma konformacijo tako, da del proteina vstavijo skozi celotno debelino membrane. Več med seboj povezanih enot proteina naredi skozi membrano nekakšen tunel oziroma poro, preko katere lahko potujejo različne snovi (Peraro in van der Goot 2016). NLP-ji pa se kljub manjši spremembi konformacije v membrano le rahlo ugreznejo. Združevanje NLP-jev nato privede do razmika lipidov, kar naredi membrano prehodno (Pirc idr. 2022).
Molekularno delovanje toksičnih NLP-jev
Škodljivci med napadom na rastlino toksične NLP-je dostavijo v neposredno bližino rastlinske celične membrane. To privede do dvojega: na eni strani do porušenja celične integritete zaradi naluknjanja membrane, kar vodi v celično smrt, na drugi strani pa povzroči aktivacijo rastlinskega imunskega sistema, ki zažene obrambne mehanizme proti invaziji patogena (Seidl in Van den Ackerveken 2019). V tem prispevku se bomo posvetili prvemu dejanju.
Kako torej pride do naluknjanja rastlinske celične membrane? Naša skupina s Kemijskega inštituta je leta 2017 s sodelavci iz tujine ugotovila, da se NLP-ji vežejo na za rastline značilne lipide GIPC oziroma na njihove glave, sestavljene iz sladkorjev, ki so izpostavljene na zunanji strani celične membrane (Lenarčič idr. 2017). Nadaljnje študije mehanizma delovanja toksičnih NLP-jev so pokazale, da je naluknjanje membrane večstopenjski proces: NLP-ji se najprej vežejo na tarčni receptor, tj. GIPC. Po vezavi se rahlo ugreznejo v membrano, v zadnjem koraku pa se povežejo v skupke oziroma agregate. Zaradi tega se lipidi razmaknejo, da nastanejo majhne heterogene pore, ki pa niso stabilne, ampak zgolj prehodne narave, kar pomeni, da nastajajo in izginjajo (Pirc idr. 2022).
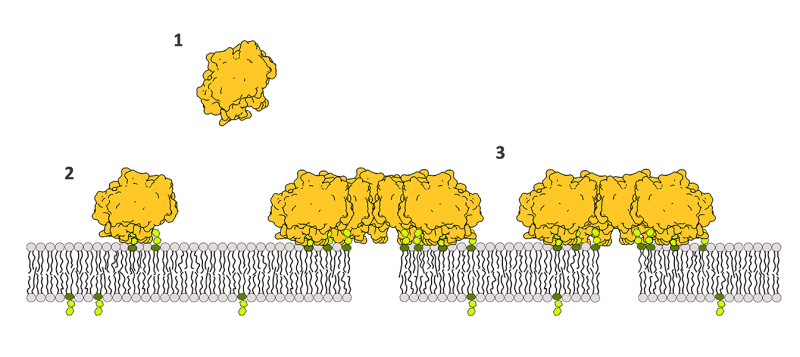
Slika 1: Mehanizem delovanja NLP-jev: (1) patogen dostavi NLP-je (v rumeni barvi) v neposredno bližino rastlinske membrane, ki med drugim vsebuje lipide GIPC (zelene sladkorne glave), (2) NLP-ji prepoznajo sladkorne glave lipidov GIPC in se nanje vežejo, temu pa sledi še (3) povezovanje NLP-jev v agregate, ki povzročijo razmak lipidov oziroma nastanek por.
Čeprav NLP-je sproščajo patogeni številnih rastlin iz vrst eno- in dvokaličnic, je bilo že zelo kmalu opaženo, da izolirani NLP-ji ne sprožijo nekroze tkiva pri vseh rastlinah (Gijzen in Nürnberger 2006). Kdor rastline malo bolje pozna in je bil pozoren, je verjetno opazil, da smo na začetku razdelka o NLP-jih pri navedbi agronomsko pomembnih tarč patogenov, ki NLP-je sproščajo, navedli le dvokaličnice. Enokaličnice so na NLP-je praviloma odporne (Lenarčič idr. 2017). Zakaj je tako? Da bomo lažje odgovorili na to vprašanje, si najprej bolje poglejmo, kdo so tarče NLP-jev oziroma lipidi GIPC.
Lipidi GIPC: najštevilnejši, izmuzljivi in nagajivi
Lipide GIPC najdemo tako pri rastlinah kot pri glivah. V rastlinski celični membrani predstavljajo nekje do 30 oziroma 40 množinskih odstotkov vseh lipidov. Upoštevajoč veliko svetovno biomaso rastlin, so lipidi GIPC najštevilčnejši lipidi na svetu (Gronnier idr. 2016). Kljub temu jih celo marsikdo, ki se ukvarja z lipidi, ne pozna (običajno so v ospredju predvsem živalski in bakterijski lipidi). Še vedno so tudi relativno slabo raziskani – kako je to mogoče?
Lipidi GIPC so sestavljeni iz značilnega hidrofobnega dela in tipične polarne glave iz sladkornih enot, ki pa nista točno definirana (Gronnier idr. 2016). Oba dela dopuščata več podobnih, a različnih molekulskih sestav. Ko govorimo o lipidih GIPC, torej ne govorimo o eni molekuli, ampak o malem morju različnih molekul, ki pa so si vseeno tako podobne, da imajo primerljive kemijske lastnosti.
Natančna sestava posameznih lipidov GIPC (z redkimi izjemami) ni znana – predvsem so za karakterizacijo težavne njihove glave, ki imajo lahko od ene do 14 sladkornih enot (Kaul in Lester 1975). Sladkorne enote so lahko različne, prav tako so lahko med seboj različno povezane.
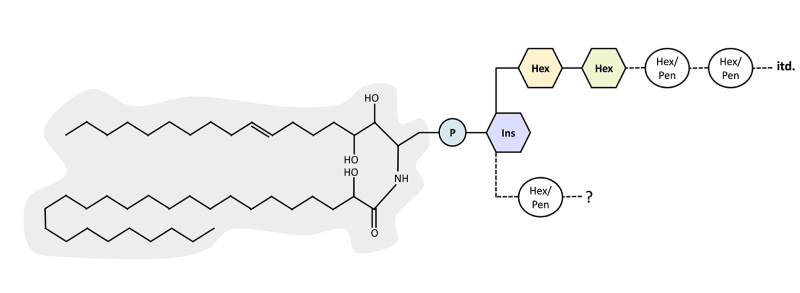
Slika 2: Molekulska sestava lipidov GIPC: hidrofobni del (označen s sivo) je prek fosfata (P) povezan z inozitolom (Ins), na inozitol pa je pripeta poljubno dolga veriga različnih sladkornih enot, tj. heksoz (Hex) in pentoz (Pen). Sladkorni verigi sta lahko tudi dve. Prikazana je ena izmed možnih sestav hidrofobnega dela – lipidni repki se lahko razlikujejo tako po dolžini kot po nasičenosti, tj. vsebnosti dvojnih vezi, in stopnji hidroksilacije, tj. številu pripetih skupin -OH.
Lipidov GIPC ne moremo kupiti, zato jih za raziskave pridobivamo z ekstrakcijo iz rastlinskega tkiva. Zaradi njihovih podobnih kemijskih lastnosti jih med postopkom med seboj ne moremo ločiti. To pomeni, da ne moremo izolirati lipida GIPC s točno določeno (željeno) molekulsko sestavo. Produkt ekstrakcije je mešanica različnih lipidov GIPC pa tudi nečistoč, ki se pri ekstrakciji obnašajo podobno kot lipidi GIPC. Zgolj delna očiščenost lipidov GIPC je eden izmed glavnih omejujočih dejavnikov pri razumevanju njihove interakcije z NLP-ji, kjer saj si pri tem želimo poznati natančno molekulsko sestavo lipida, ki ga NLP prepozna.
Grobo očiščene pripravke lipidov GIPC bi v teoriji lahko nadaljnje ločili z bolj specifičnimi tehnikami. Vendar to v praksi ni tako enostavno, saj so že za pridobitev majhne količine ekstrakta potrebne velike količine vhodnega rastlinskega materiala. Posameznih zvrsti lipidov GIPC pa je v ekstraktu zelo malo.
Sestava različnih molekul lipidov GIPC se med rastlinami razlikuje, a v splošnem velja, da je v rastlinski membrani dvokaličnic praviloma največ takih lipidov GIPC, katerih glava vsebuje dve sladkorni enoti (GIPC serije A), pri enokaličnicah pa prevladujejo glave s tremi sladkornimi enotami (GIPC serije B). V manjši količini so pri vseh rastlinah prisotni tudi lipidi GIPC z eno sladkorno enoto (GIPC serije 0), pa tudi tisti s štirimi ali več (GIPC serij C, D, E itd.) (Gronnier idr. 2016).
Interakcija NLP-jev z lipidi GIPC
Omenili smo že, da so dvokaličnice praviloma dovzetne za napade toksičnih NLP-jev, enokaličnice pa ne. Prav tako smo v prejšnjem razdelku navedli, da pri dvokaličnicah prevladujejo lipidi GIPC serije A, pri enokaličnicah pa lipidi GIPC serije B. Naši raziskovalci s Kemijskega inštituta so s sodelavci iz tujine ugotovili, da NLP-ji vsebujejo posebno vezavno mesto za končno sladkorno enoto lipidov GIPC. To je glavno mesto interakcije, poleg tega pa je najverjetneje prisotnih še več stranskih, manj specifičnih interakcij NLP-ja z bližnjimi lipidi GIPC (Lenarčič idr. 2017; Pirc idr. 2022). Iz tega bi lahko enostavno sklepali, da NLP-ji ne razlikujejo med posameznimi serijami GIPC.
Študije vezave so res pokazale, da se NLP-ji vežejo tako na lipide GIPC iz serije A kot na lipide GIPC iz serije B, vendar membrana postane prepustna le v primeru, da vsebuje pretežno lipide GIPC serije A. To je v skladu z opažanji, da izolirani NLP-ji običajno sprožijo nekrozo pri dvokaličnicah, pri enokaličnicah pa ne. Sledila je logična hipoteza, da so sladkorne glave lipidov GIPC serije A dovolj kratke, da lahko NLP po vezavi dostopa do membrane, v primeru vezave na lipide GIPC iz serije B pa to ni mogoče (Lenarčič idr. 2017).
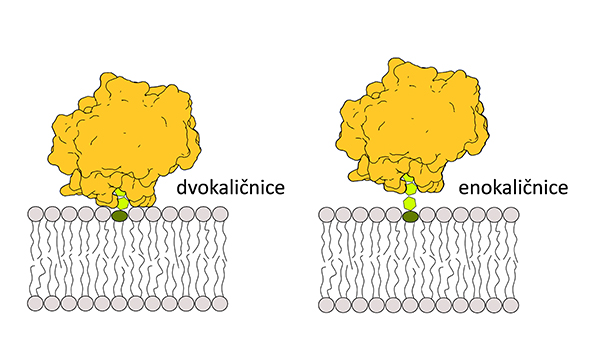
Slika 3: Vezava NLP-ja (v rumeni barvi) na lipidno glavo GIPC (z zeleno) z dvema (levo) oziroma tremi sladkornimi enotami (desno). Razdalja do membrane je pri treh sladkornih enotah prevelika, da bi jo NLP dosegel.
Skok v neznano – katera tarča je prava?
Mogoče se zdaj zdi, da je vse sila enostavno in da je interakcija med NLP-ji in lipidi GIPC dobro pojasnjena, vendar to ne drži. V zadnjem času se pojavljajo navedbe, da lahko NLP-ji sprožijo nekrozo tudi pri nekaterih enokaličnicah, pri čemer njihova toksičnost ne sovpada z večjo količino lipidov GIPC serije A v membrani (Steentjes idr. 2022). Da lahko NLP-ji naluknjajo membrano enokaličnice, je bilo pokazano tudi na protoplastih, tj. rastlinskih celicah brez celične stene (Lin idr. 2023).
Razlaga bi se lahko skrivala v tem, da je delitev lipidov GIPC na serije (glede na število sladkornih enot v njihovi polarni glavi) preveč enostavna, saj ne upošteva vseh dejavnikov, ki bi lahko vplivali na to, kako NLP-ji prepoznavajo lipide GIPC. Polarne glave znotraj posamezne serije GIPC se namreč razlikujejo, in sicer po tem, katere sladkorne enote vsebujejo in kako so te med seboj povezane. Je potrebna točno določena kombinacija sladkornih enot? Je pomembno, kako so med seboj povezane? Je tarča NLP-jev ena specifična molekula GIPC ali pa je tarč več? Obstajajo še drugi dejavniki?
Ker je v ekstraktih, ki se navadno uporabljajo za raziskovanje interakcije NLP-jev z lipidi GIPC, veliko različnih molekul GIPC pa tudi nečistoč, z njimi ne moremo identificirati posameznih molekul GIPC kot tarč NLP-jev. Zato smo se na Kemijskem inštitutu s sodelavci pred kratkim lotili nadaljnjega čiščenja lipidov GIPC iz ekstraktov in natančne karakterizacije njihovih sladkornih glav. Pri delu z NLP-ji nas zanimajo le sladkorne glave lipidov GIPC, zato smo pri ločevanju zvrsti zanemarili razlike v njihovih hidrofobnih delih. Z izolati, ki vsebujejo lipide GIPC z definiranimi enakimi sladkornimi glavami, želimo natančno raziskati, kako sestava lipidne glave vpliva na različne stopnje delovanja proteinov NLP.
Ko bi le bilo tako enostavno …
Poleg sestave sladkornih glav lipidov GIPC lahko na nastanek por vplivajo tudi drugi dejavniki. Znana je npr. vloga sterolov, ki spodbujajo povezovanje molekul GIPC v nekakšne otočke v membrani oziroma lipidne rafte. Steroli olajšajo tvorbo por (Pirc idr. 2022), ni pa še znano, zakaj. Poročalo se je tudi že o oligomerizacijskem faktorju, tj. proteinu, ki spodbuja agregacijo NLP-jev po njihovi vezavi na membrano in s tem olajša njeno naluknjanje (Chen idr. 2021). Na drugi strani je bil pred kratkim odkrit rastlinski protein, ki zavira delovanje NLP-jev (Wang idr. 2023). Delovanje obeh proteinskih dejavnikov na molekularnem nivoju še ni dobro pojasnjeno, zato ju v zgodbo o napadu NLP-jev na rastline zaenkrat še ne moremo zares umestiti. Vidimo pa, kako pomembno je, da pri preučevanju porotvorne aktivnosti NLP-jev uporabljamo kontrolirane modelne sisteme, pri katerih se takšnim spremenljivkam izognemo.
Nepojasnjeno ostaja tudi to, ali so mehanizmi delovanja različnih toksičnih NLP-jev enaki oziroma kakšne so razlike. Bi lahko natančnejše znanje privedlo do razvoja pesticidov, ki bi bili zelo specifični, hkrati pa učinkoviti proti številnim fitopatogenom?
Kot vidimo, nas na področju razumevanja delovanja NLP-jev in njihove interakcije z lipidi GIPC čaka še veliko dela. Vseeno pa bomo tokrat zaključili s precej enostavno ugotovitvijo: NLP-ji z neznanimi letečimi predmeti nimajo skupne le kratice, temveč tudi to, da o njih še marsičesa ne vemo in da so za raziskovanje vse prej kot enostavni.



